![]()
| 横川浩治(香川県多度津町)・ |
| 中井克樹(琵琶湖博物館)・藤田建太郎(滋賀県立大学) |
|
|
オオクチバス Micropterus salmoides は北米原産の淡水魚で、これには北米大陸の中北部に分布するノーザンバス M. salmoides salmoides と、主にフロリダ半島に分布するフロリダバス M. salmoides floridanus の2亜種の存在が認められているが、最近の形態的および遺伝的研究によりフロリダバスは独立種 Micropterus floridanus として扱うべきとする見解もある(Kassler et al., 2002)。
ノーザンバスとフロリダバスを形態的に区別する決定的な形質は側線有孔鱗数であり、ノーザンバスでは68枚以下であるのに対して、フロリダバスでは70枚以上であるとされている(Tomelleri and Eberle, 1990)。その他の特徴として、フロリダバスはノーザンバスよりも成長度が卓越しているとされ、全長70cmを越えるものも珍しくないと言われている。さらに、これら2(亜)種は容易に交雑を起こすこともよく知られている(Pelzman, 1980; Tomelleri and Eberle, 1990)。北川ら(2000)は奈良県の池原貯水池のオオクチバスのミトコンドリアDNAを調べ、集団内にフロリダバスの遺伝子が混入していることを発見したが、日本国内でフロリダバス系のオオクチバスの生息が確認されているのは現在のところ池原貯水池だけである。
かつて、Yokogawa(1998)は全国各地のオオクチバスを形態的および遺伝的に調べ、琵琶湖産のオオクチバスもノーザンバスであることを確認した。しかし最近、琵琶湖にフロリダバスが大規模に侵入したことが明らかとなったので報告する。
| Table 1. 調べた標本のデータ |
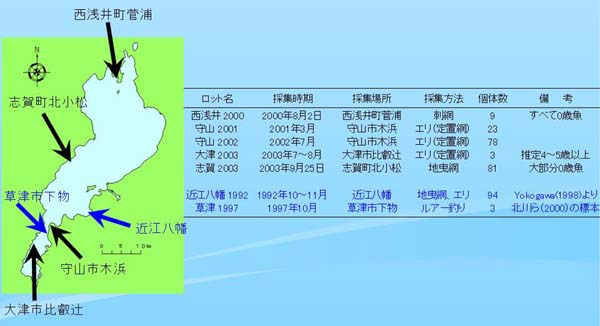 |
今回調べた標本に関するデータを Table 1 に示す。標本は2000年から2003年にかけて採集し、琵琶湖の最北にある西浅井、南湖と北湖の東岸にまたがる守山、大津(南湖西岸)、および北湖西岸の志賀町からサンプルを得た。大津2003の標本群は3個体と少ないが、すべて全長50㎝を超える大型の個体で、年級としては西浅井2000の個体群よりも以前の発生群と推定される。
今回得られた標本魚については Yokogawa(1998)に従い、水平式デンプンゲル電気泳動法によりアイソザイムを調べた。
なお参考データとして、Yokogawa(1998)による1992年に近江八幡で得られた94個体のアイソザイムデータと、北川ら(2000)が1997年に草津市下物で採集してミトコンドリアDNAを調べた個体を譲り受けて調べたアイソザイムデータも合わせて用いた(Table 1)。
| 遺伝子座 | 遺伝子1 | 近江八幡1992 | 草津1997 | 西浅井2000 | 守山2001 | 守山2002 | 大津2003 | 志賀2003 | |
| AAT-1* | 230 | D | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| 200 | C | 0.000 | 0.000 | 0.444 | 0.250 | 0.625 | 0.500 | 0.389 | |
| 145 | B | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 100 | A | 1.000 | 1.000 | 0.556 | 0.750 | 0.375 | 0.500 | 0.611 | |
| IDHP-1* | 150 | C | 0.000 | 0.000 | 0.000 | 0.000 | 0.006 | 0.000 | 0.000 |
| 120 | B | 0.000 | 0.000 | 0.556 | 0.413 | 0.596 | 0.500 | 0.475 | |
| 100 | A | 1.000 | 1.000 | 0.444 | 0.587 | 0.397 | 0.500 | 0.525 | |
| MDH-1* | 140 | B | 0.005 | 0.000 | 0.444 | 0.239 | 0.558 | 0.500 | 0.469 |
| 100 | A | 0.995 | 1.000 | 0.556 | 0.761 | 0.442 | 0.500 | 0.531 | |
| SOD* | 100 | A | 1.000 | 1.000 | 0.889 | 0.826 | 0.870 | 1.000 | 0.889 |
| 45 | B | 0.000 | 0.000 | 0.111 | 0.174 | 0.130 | 0.000 | 0.111 | |
| 1 アルファベットの前の数字は、ノーザンバスにおける主遺伝子(A)に対する相対的なバンドの移動度を百分率で表わしたもの. 水色はノーザンバスに固有の対立遺伝子、黄色はフロリダバスに固有の対立遺伝子. |
|||||||||
ノーザンバスとフロリダバスの遺伝的特徴については原産地の北米では非常に詳しく調べられており、Table 2 に示す四つのアイソザイム系遺伝子座で両者を識別できることが報告されている(Philipp et al., 1983)。
そのうちのひとつである Idh-B 遺伝子座(肝臓におけるイソクエン酸脱水素酵素)の遺伝子組成によれば、フロリダ半島を中心に泳動速度の速い B3 遺伝子を持つフロリダバスの集団が分布していて、北米大陸の中部以北は泳動速度の遅い B1 遺伝子で特徴づけられるノーザンバス、その間には両者の Intergrade zone が存在することが明瞭に示されている(Philipp et al., 1983)。
本研究では Yokogawa(1998)に従って Philipp et al.(1983)の Idh-B 遺伝子座を IDHP-1* 遺伝子座とし、ノーザンバスに固有のB1遺伝子をA遺伝子、フロリダバスに固有のB3遺伝子をB遺伝子と表記する。
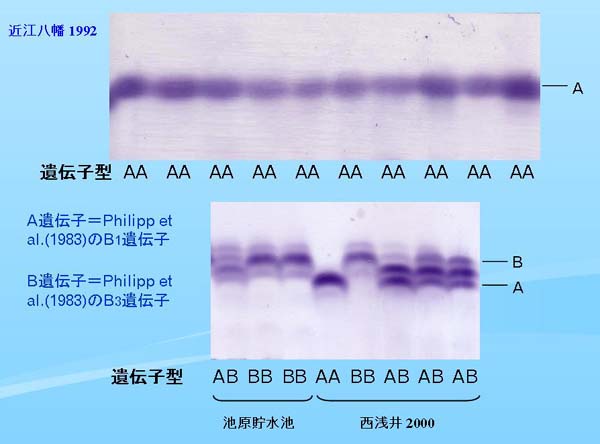 |
| Fig.1. 肝臓におけるイソクエン酸脱水素酵素(IDHP)の泳動像 |
琵琶湖産オオクチバスの肝臓における IDHP の泳動像を Fig. 1 に示す。上段は近江八幡1992の泳動像だが、すべての個体で全く同一のホモ型で、他の地域のオオクチバスのとの泳動像の比較や形態的特徴から総合的に判断して、これらはすべてノーザンバス固有のAA型のホモ個体と判断された(Yokogawa, 1998)。
Fig. 1 の下段は西浅井2000の個体と、北川ら(2000)によってフロリダバスが遺伝的に混入していることが確認された池原貯水池の個体を並べて泳動を行なった結果である。西浅井2000では以前の琵琶湖ではみられなかった変異個体が多く出現し、変異バンドの位置は池原貯水池のB遺伝子と一致することから、西浅井で出現する変異はフロリダバスに固有のB遺伝子であると判断された。
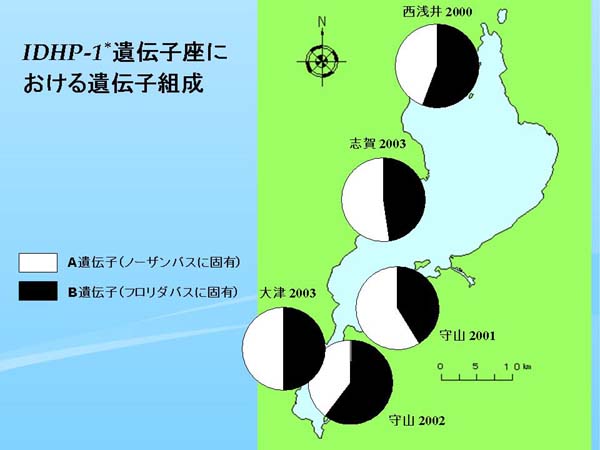 |
| Fig. 2. 各標本群のIDHP-1* 遺伝子座における遺伝子組成 |
琵琶湖におけるこの IDHP-1* 遺伝子座の遺伝子組成を円グラフにして地図上にプロットした(Fig. 2)。琵琶湖で最も古いアイソザイムデータである近江八幡1992、および草津1997ではすべてノーザンバス固有のA遺伝子で占められていたが(Table 2)、西浅井 2000ではフロリダバス固有のB遺伝子が出現し、全体の半分以上の頻度を占めた。さらに、守山2001、守山2002、大津2003、志賀2003も類似した傾向を示し、近年の琵琶湖ではフロリダバス固有の遺伝子が概ね全体の半分を占めると言えよう(Table 2、Fig. 2)。
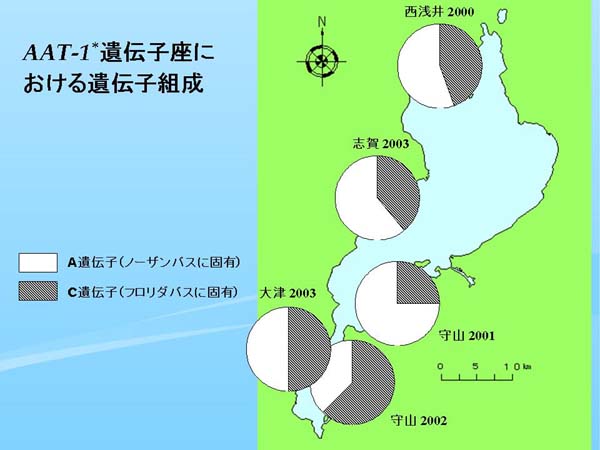 |
| Fig. 3. 各標本群のAAT-1* 遺伝子座における遺伝子組成 |
ノーザンバスとフロリダバスを識別できる別のマーカー座である AAT-1* 遺伝子座(肝臓におけるアスパラギン酸アミノ転移酵素、Philipp et al., 1983では Aat-B locus)においては、原産地の北米ではノーザンバスの集団は泳動速度の遅いA遺伝子とB遺伝子のみの構成であり、フロリダバスの方は泳動速度の速いC遺伝子とD遺伝子で占められるという形で対立遺伝子が置換している(Philipp et al., 1983)。
琵琶湖における AAT-1* 遺伝子座の遺伝子組成を円グラフにして地図上にプロットした(Fig. 3)。琵琶湖で最も古いデータである近江八幡1992、および草津1997ではすべてノーザンバス固有のA遺伝子で占められていたが(Table 2)、西浅井2000ではフロリダバス固有のC遺伝子が出現し、全体の半分近くの頻度を占めた。一方、守山2001ではフロリダバスの遺伝子頻度は4分の1だが、守山2002では3分の2に近く、大津2003と志賀2003ではC遺伝子がほぼ半分を占めた(Table 2、Fig. 3)。
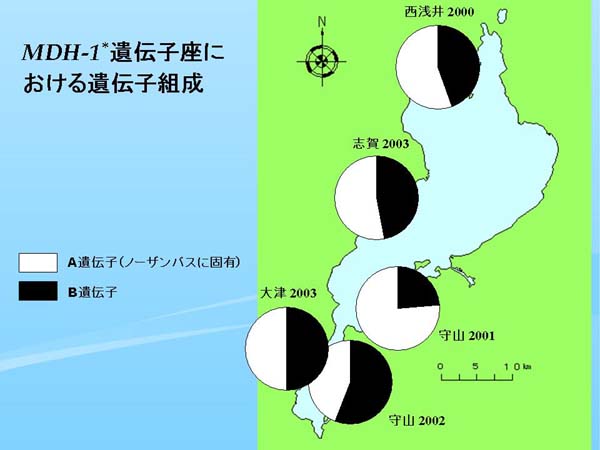 |
| Fig. 4. 各標本群のMDH-1* 遺伝子座における遺伝子組成 |
MDH-1* 遺伝子座(筋肉におけるリンゴ酸脱水素酵素、Philipp et al., 1983では Mdh-B locus)では、原産地の北米ではフロリダバスはすべての集団で泳動速度の速いB遺伝子で固定されている。一方ノーザンバスの方は、固有のA遺伝子で固定される場合もあるがB遺伝子を含む集団が多く、この遺伝子座においては対立遺伝子が完全置換はしていない。しかし、B遺伝子はフロリダバスを特徴づけるマーカーと言える(Philipp et al., 1983)。
琵琶湖における MDH-1* 遺伝子座の遺伝子組成を円グラフにして地図上にプロットした(Fig. 4)。近江八幡1992ではB遺伝子がわずかにみられたが(Table 2)、これは、調べた94個体中でAB型の個体が1個体だけ出現したためである。一方、草津1997ではノーザンバス固有のA遺伝子で占められており(Table 2)、このような遺伝的状況にB遺伝子しか持たないフロリダバスが混入すれば明瞭に判明するはずである。
そこで、西浅井2000ではB遺伝子が全体の半分近くの頻度を占め、守山2001ではB遺伝子の頻度は4分の1程度だが、守山2002では3分の2に近く、大津2003と志賀2003ではC遺伝子がほぼ半分を占めた(Table 2、Fig. 4)。MDH-1* 遺伝子座における遺伝子頻度の分布図は AAT-1* 遺伝子座の場合(Fig. 3)と非常によく似た結果となり、フロリダバスの遺伝子が大規模に混入したことが示唆された。
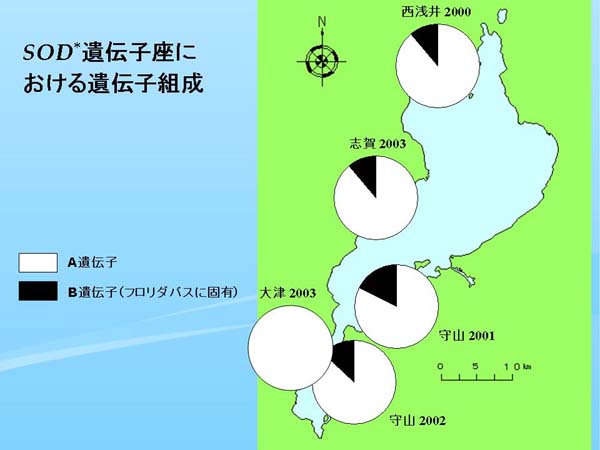 |
| Fig. 5. 各標本群のSOD* 遺伝子座における遺伝子組成 |
SOD* 遺伝子座(肝臓におけるスーパーオキサイドディスムターゼ、Philipp et al., 1983では SOD-A locus)では、原産地の北米ではノーザンバスはすべての集団で泳動速度の速いA遺伝子で固定されている。一方フロリダバスの方は泳動速度の遅い固有のB遺伝子が出現するのが特徴だが、完全置換はしていない。しかし、B遺伝子はフロリダバスを特徴づけるマーカーと言え、B遺伝子が出現すればフロリダバスが混入している証拠となる(Philipp et al., 1983)。
琵琶湖における SOD* 遺伝子座の遺伝子組成を円グラフにして地図上にプロットした(Fig. 5)。最も古いデータである近江八幡1992、および草津1997ではすべてノーザンバス固有のA遺伝子で占められていたが(Table 2)、2000年以降の個体群では大津2003を除いてすべてフロリダバス固有のB遺伝子が出現した(Fig. 5)。大津2003でB遺伝子が出現しなかったのは調べた個体数が3個体と少ないためと思われ(Table 1)、これらのことは近年琵琶湖にフロリダバスが侵入したという明確な証拠と言える。
|
||
|
||
|
||
|
ここで、マーカー座における遺伝子型による個体ごとの類型分類を試み、Table 3 の定義によって四つの Type に類別した。
Type 2 については、MDH-1* 遺伝子座は原産地の北米では完全置換ではないが、琵琶湖ではフロリダバスが入る以前はB遺伝子はほとんどなかったことから(Table 2)完全置換と見なし、③の条件が成立する。
| 個体番号 | B-1 | B-2 | B-3 | B-4 | B-5 | B-6 | B-7 | B-8 | B-9 |
| AAT-1* | CC | AA | AC | AC | AC | AC | AC | AC | AC |
| IDHP-1* | BB | AA | AB | BB | AA | BB | AB | AB | AB |
| MDH-1* | AB | AA | AB | AB | BB | AB | AB | AA | AB |
| SOD* | AA | AA | AA | AA | AA | AA | AA | AB | AB |
| Type | 4 | 1 | 3 | 4 | 4 | 4 | 3 | 4 | 3 |
| 側線鱗数 | 71 | 63 | 69 | 66 | 68 | 67 | 69 | 64 | 69 |
類型判別の具体例として西浅井2000における類型判別の結果を Table 4 に示す。黄色の部分が Type 1、水色が Type 3、無印が Type 4 で、ここでは Type 2 は出現しなかった。
ここで、Typeと側線鱗数に着目すると、Type 3 の雑種 F1 と判定される3個体では側線鱗数はすべて69枚で、68枚以下であるノーザンパスと70枚以上であるフロリダバス(Tomelleri and Eberle, 1990)のちょうど中間の値となっている。
また Type 4 でも B-1 の個体のように側線鱗数が70枚を超えるものもあり、この個体は遺伝子型からするとおそらく F1 と純粋なフロリダバスの戻し交雑個体と思われるが、このように、雑種化が進行すれば形態形質による判別は非常に難しいことが示唆される(Table 4)。
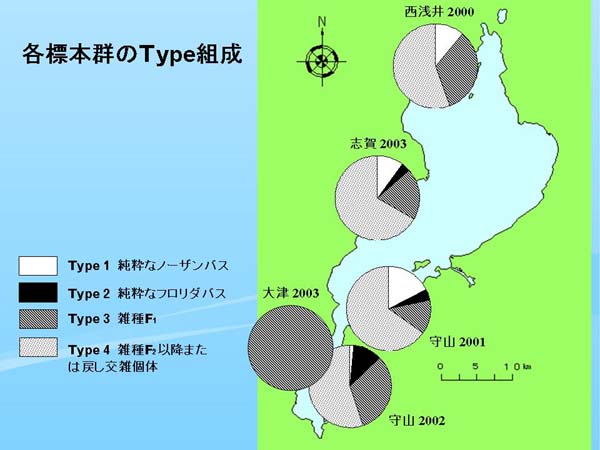 |
| Fig.6. 各標本群の Type 組成 |
ここで、標本群ごとの Type 組成を円グラフにして地図上にプロットした(Fig. 6)。西浅井2000では、Type 3 と Type 4 が主体の組成であり、守山2001では Type 4 が全体の3分の2くらいを占めるが、Type 1 の純粋なノーザンバスもやや多かった。守山2002では西浅井と類似した組成だが、Type 2 の純粋なフロリダバスがやや多かった。大津2003では3個体とも雑種 F1 であったが、前述のように大津の標本群が今回調べたうちでいちばん古い年級と思われ、これらの個体はあるいは琵琶湖にフロリダバスが侵入した直後に発生した雑種である可能性も考えられる。また、志賀2003では守山2001とよく似た組成であった。
全体に、Type 4 の占める割合が高く、湖内のどの場所でも雑種化がかなり進行しつつあるもの思われた。
| 近江八幡1992 | 草津1997 | 西浅井2000 | 守山2001 | 守山2002 | 大津2003 | 志賀2003 | ||
| 平均対立遺伝子数 | 1.050 | 1.000 | 1.320 | 1.360 | 1.440 | 1.208 | 1.308 | |
| P* | 0.000 | 0.000 | 0.360 | 0.200 | 0.200 | 0.125 | 0.192 | |
| P | 0.050 | 0.000 | 0.000 | 0.120 | 0.240 | 0.000 | 0.154 | |
| P+P* | 0.050 | 0.000 | 0.360 | 0.320 | 0.440 | 0.125 | 0.346 | |
| 平均ヘテロ接合体率 | Ho | 0.001 | 0.000 | 0.111 | 0.077 | 0.091 | 0.125 | 0.062 |
| He | 0.001 | 0.000 | 0.093 | 0.069 | 0.074 | 0.063 | 0.071 | |
| Ho/He | 1.005 | - | 1.194 | 1.125 | 1.225 | 2.000 | 0.875 | |
| P* : 最大対立遺伝子頻度が0.95以下の遺伝子座の割合 P : 最大対立遺伝子頻度が0.95以上の遺伝子座の割合 Ho : 平均ヘテロ接合体率の観察値 He : 平均ヘテロ接合体率の期待値 |
||||||||
今回調べた標本群の遺伝的諸特性値の一覧を Table 5 に示す。2000年以降の標本群ではそれ以前のものに比べて平均対立遺伝子数や平均ヘテロ接合体率の値がかなり高く、遺伝的多様性が高いことがわかる。これは明らかにフロリダバスの遺伝子が大量に入ったためである。
平均ヘテロ接合体率の観察値と期待値の比(Ho/He)は、志賀2003を除いてすべて1を上回りヘテロ過剰の傾向を示した。これは、どの個体群内にも交雑個体(Type 3, 4 )が多いこと(Fig. 6)によるものと思われる。
| IDHP-1* | 遺伝子型 | AA | BB | AB | |
| 観察値 | 23 | 19 | 39 | ||
| 期待値 | 22.3 | 18.3 | 40.4 | ||
| 0.097 | |||||
| MDH-1* | 遺伝子型 | AA | BB | AB | |
| 観察値 | 24 | 19 | 38 | ||
| 期待値 | 22.8 | 17.8 | 40.3 | ||
| 0.274 |
今回調べたすべての標本群のすべての多型的遺伝子座について、Hardy-Weinberg の遺伝平衡への適合性を検定によって調べた。その一例として志賀2003における計算を Table 6 に示す。この例では両遺伝子座とも各遺伝子型個体数の観察値と期待値は非常によく一致して有意差は認められなかったが、これと同様にすべての標本群の多型座において遺伝平衡からのずれは認められなかった。このことから琵琶湖内では既にノーザンバスとフロリダバスの交雑がかなり進行していることが示唆され、これは、前述のようにどの個体群にも Type 4 の個体が多いことからも支持される(Fig. 6)。
 |
| Fig. 7 琵琶湖産オオクチバスの形態の推移 |
琵琶湖産オオクチバスの形態の推移を Fig. 7 に示す。上段は近江八幡1992の標本群中の1個体で純粋なノーザンバスであるが、体高は低くて体型は細身で、体側の縦帯は明瞭である。
一方下段は西浅井2000の標本群中の1個体で、Type 4 の交雑が進んだ個体である。上の個体とほとんど同じ全長15㎝程度のものだが、体高が高くまた体側の縦帯は全く不明瞭で、以前の純粋なノーザンバスとはかなり概観が異なる。
現在琵琶湖に生息するオオクチバスの多くはこのような個体であり、いずれすべてがこのようになるものと予想される。
まとめ
引用文献
- 琵琶湖には近年、フロリダバスが大量に侵入し、交雑によってノーザンバスあるいはフロリダバスの遺伝子が選択的に残存するメカニズムがないとすれば、侵入したフロリダバスは従来生息していたノーザンバスに匹敵する数量であったと推定される。
- 現在、琵琶湖内ではノーザンバスとフロリダバスの交雑が大規模に進行しつつあり、近い将来にはすべて交雑個体となるであろう。
- このような現象が自然に発生する可能性はあり得ず、人為的な大規模放流があったものと推定される。
- Kassler, T. W., J. B. Kopperman, T. J. Near, C. B. Dillman, J M. Levengood, D. L. Swoffoord, J. L. VanOrman, J. E. Claussen and D. P. Philipp. 2002. Molecular and morphological analyses of the black basses: implications for taxonomy and conservation. Pages 291-322 in D. P. Philipp and M. S. Ridgway, eds. Back bass: ecology, conservation, and management. Am. Fish. Soc., Bethesda, US.
- 北川忠夫・沖田智明・伴野雄次・杉山俊介・岡崎登志夫・吉岡 基・柏木正章.奈良県池原貯水池から検出されたフロリダバス Micropterus salmoides floridanus 由来のミトコンドリアDNA.日本水産学会誌,66(5): 805-811.
- Pelzman, R. J. 1980. Impact of Florida largemouth bass, Micropterus salmoides floridanus, introductions at selected northern California waters with a discussion of the use of meristics for detecting introgression and for classifying individual fish of intergraded populations. Calif. Fish and Game, 66(3): 133-162.
- Philipp, D. P., W. F. Childers and G. S. Whitt. 1983. A biochemical genetic evaluation of the northern and Florida subspecies of largemouth bass. Trans. Am. Fish. Soc., 112(1): 1-20.
- Tomelleri, J. R. and M. E. Eberle. 1990. Fishes of the central United States. Univ. Press Kansas, Laurence, US. xv+226 pp.
- Yokogawa, K. 1998. Morphological and genetic structures of largemouth bass Micropterus salmoides in Japanese fresh waters. Suisanzoshoku, 46(3): 321-332.
* このウエブサイトは、2004年度日本魚類学会年会(於 琉球大学西原キャンパス)で著者らのひとり横川が口頭発表を行なった内容を公開するものです。すべてのコンテンツの無断転載を禁じます。
* なお本研究の内容は、以下の通り日本水産増殖学会誌に原著論文として印刷されています。以下のリンクをクリックすれば原著論文をご覧になれます。
Yokogawa, K., N. Nakai and F. Fujita. 2005. Mass introduction of Florida bass Micropterus floridanus into Lake Biwa, Japan, suggested by recent dramatic genomic change. Aquacult. Sci., 53(2): 145-155.